





- Characteristics of Microbial Community Enzyme Activity and Substrate Availability of Damaged Soil
Ji Seul Kim1·Gyo-Cheol Jeong2·Myoung Hyeon Cho3·Eun Young Lee4*
1Doctoral Program, Department of Environmental Engineering, The University of Suwon, Korea
2Professor, Department of Earth and Environmental Sciences, Andong National University, Korea
3Director, NEXUS Environmental Design Center, Korea
4The University of Suwon, Department of Environmental and Energy Engineering, Professor, Korea- 훼손 토양의 미생물군집 효소 활성과 기질 이용성 특성
김지슬1·정교철2·조명현3·이은영4*
1수원대학교 환경공학과 박사과정, 2안동대학교 환경공학과, 명예교수,
3넥서스 환경디자인연구원 ㈜, 원장, 4수원대학교 환경에너지공학과, 교수This article is an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
The effect of soil damage on the physicochemical characteristics and activity of the soil microbial community is not well known. This study investigates this relationship by analyzing 11 soil samples collected from various points of soil damage across Gyeonggi-do. Soil damage resulted from forest fires, landslides, and development areas, with their impacts most severe on the topsoil layer (0-30 cm). Dehydrogenase and b-glucosidase activities were notably higher at locations damaged by forest fires compared to other sites. While enzyme activities in soils influenced by landslides and development areas were relatively low, sites with a pollution history exhibited elevated dehydrogenase activity, likely due to past microbial response to the pollution. Additionally, an assessment of carbon substrate usability by soil microorganisms indicated higher substrate availability in areas impacted by forest fires, contrasting with lower availability in landslide and development sites. Statistical analysis revealed a positive correlation between organic content of sand and clay and microbial activity. These findings provide valuable insights into soil damage and associated restoration research, as well as management strategies.
Keywords: Damaged soil, Dehydrogenase, b-glucosidase, Soil microbial community, Biological indicator
지구의 표면을 덮고 있는 토양은 생물의 생산, 배양과 분해, 양분 및 수분의 저장 등 다양한 기능을 하는 중요한 환경매개체이다. 그러나 여러 가지 원인으로 토양의 훼손이 진행되고 있는데, 태풍, 홍수, 쓰나미, 화산, 산불과 같은 자연재해 및 기후변화 등에 의한 자연적인 요인이 있으며, 인위적 요인으로는 토지 개발과, 산림 벌채, 산불, 불합리한 농업관리, 광업 활동 및 유해물질 배출에 따른 오염에 의한 훼손 등이 있다. 전 국토가 도시화 되는 과정에서 발생한 이러한 훼손은 도시지역 내의 작은 자연환경 면적에서 격리된 채 외부의 교란 요인에 대하여 적응하기도 하나, 기후환경의 변화에 대응하지 못하고 쉽게 쇠퇴하기도 한다. 이렇게 훼손된 지역의 경우 토양이 우리에게 제공하는 여러 가지 직·간접적인 가치를 훼손하게 되는데, 생물 다양성의 훼손, 오염물질의 자연적 정화과정의 중단, 자연재해 및 외부 변화에 대한 완충작용 등을 할 수 없게 된다.
국내에서는 최근 10년간 5,368건의 산불이 발생하였으며, 35,597 ha의 피해면적이 발생하였다. 산불은 동·식물의 서식지를 파괴하고, 생물 다양성을 감소시키며, 토양미생물을 감소시키며(Zhang et al., 2020), 이차적으로 홍수가 발생되기 쉬우며 표토 유실을 증가시킨다. 화재는 불이 닿는 표토와 심토의 토양 입단구조 변형을 일으키며, 토양의 물리, 화학적 조직에 변화를 일으킨다. 특히 토양 조성과 광물 구성을 변화시킨다(Kim and Choo, 2023). 화재 온도가 특정 온도 이상 올라가면 광물 구조의 변화가 열적 응력을 유발하고, 양이온교환능력(CEC)이 감소한다(Kim and Choo, 2023; Mataix-Solera et al., 2011). 양이온 교환 능력은 토양이 입도가 작아질수록 증가하고, 조립질이나 사질 입자에서 감소하는데, 산불이 발생되면 광물 구조가 열적 응력으로 입자 사이즈가 변화되어 화학적 변화를 초래하는 것으로 보여진다. 산불 훼손지에서는 점토광물의 일종인 일라이트 / 버미큘라이트 혼합층상 구조 광물을 관찰할 수 있으나, 미 발생지에서 관찰되지 않은 산불로 인한 광물조성의 변화도 보고되었다(Kim and Choo, 2023). 또한, 점토광물이 함수비 증가 시 팽창과정에서 응력을 발생시켜 지반구조의 안정성에 문제를 야기할 수 있다고 한다. 이와 같이 다양한 토양 훼손의 원인과 특성을 연구하는 것은 훼손이 진행되는 것을 방지하고 복원을 계획하는 것의 시작이 될 수 있다.
현재까지도 토양의 훼손이 토양의 분해를 가져오고 토양 미생물군집과 토양의 다양한 기능에 영향을 미치는 기작에 대하여는 잘 알려져 있지 않다. 또한, 토양의 특성과 물리·화학적 성질이 토양미생물의 활성에 미치는 영향도 많은 연구가 이루어지지 않은 상태이다. 지금까지는 산불 혹은 훼손지의 환경적 변화로 인한 토양의 물리적 특성과 화학적 특성에 관한 연구가 진행되었으나, 토양의 건전성 및 회복을 판단하기 위해서 물리적 특성과 생물학적 지표와의 종합적 판단이 필요하다. 훼손 지역의 물리·화학적 특성은 큰 차이점을 보이지 않는다. 그러나 아주 미세한 물리·화학적 변화에 대하여 생물학적 활성은 매우 민감한 변화를 보여준다. 그러므로, 생물학적 활성을 분석한 결과는 각 훼손지역에서 복원의 진행되는 과정을 모니터링하는데 유용한 수단이 될 수 있다.
따라서 본 연구에서는 다양한 토양 훼손 지역 중 산불 훼손지, 산사태 훼손지, 도시개발지역의 훼손지를 선정하여 각 지역의 표토를 채취하여 물리·화학적인 특성과 토양미생물의 효소 활성을 알아보고 각 요인의 상관관계를 분석하였다.
본 연구에서 살펴본 생물학적 지표 인자로는 토양 미생물군집의 기질 이용성, 효소 활성 등을 알아보았다. 토양 미생물은 환경에 영향을 미쳐 토양에 서식하는 식물이 자랄 수 있게 하고 번성하게 해준다. 새롭게 토양이 형성되게 되면 광합성 미생물이 토양에 정착하게 되고, 질소, 탄소, 인의 순환을 하고, 토양에 영양분을 제공하는 역할을 하게 된다. 또한, polysaccharide 및 glycoprotein을 분비하여 토양입자의 표면을 둘러싸고, 안정적인 미세구조를 형성하여 토양 구조를 개선하게 된다(Hoorman et al., 2011). 토양의 훼손이 적을수록 토양에 자생하는 식물의 다양성이 커지며, 토양의 먹이그물이 보다 균형적이며 다양해져서 고등식물에 더욱 적합한 환경이 된다. 토양 효소는 토양의 생물학적 특성 및 토양 질 평가지표로 이용되는데(Alkortal et al, 2003), 효소의 활성은 온도, 습도, 토양 양분 등의 비생물적 요인 및 토양 미생물의 밀도, 다양성 등 생물학적 요인에도 영향을 받는다(Joa et al., 2009). 다양한 물리·화학적 환경 조건은 토양의 효소 활성에 영향을 미치기 때문에 특정 지역의 토양 효소 활성을 측정함으로써 해당 지역의 토양 건전성을 평가할 수 있다(Langer and Günther, 2001). 그 중, b-글루코시다아제는 토양 내 섬유소를 분해하는 효소로 토양 탄소순환과정에 기여하고 있다. 일반적으로는 토양 유기물 함량 및 바이오매스 함량이 증가할수록 효소 활성이 증가한다(Aranda et al., 2015). 효소 활성은 pH 조건에 따라 달라질 수 있으며, 산성에서 활성이 높은 것으로 알려져 있다(Puissant et al., 2019; Wade et al., 2021). 탈수소효소는 토양 유기물의 생화학적 산화환원에 필수적인 역할을 하며, b-글루코시다아제와 마찬가지로 유기물 함량이 높아질수록 효소 활성이 높아진다(Chun et al., 2021). 그러나, 이러한 효소의 활성은 유기물 함량 외에 토성, 점토 함량, 미생물 생체량 및 기타 오염물질 등에도 영향을 받는 것으로 보고되어있다(Acosta-Martinez et al., 2007; Aponte at al., 2020; Kim et al., 2023)
최근까지 연구자들은 특정 환경 내에서 유기물, 지역, 화학 물질 등이 미생물 효소 활성에 미치는 영향을 분석함으로써 토양 건전성과 토양 미생물 군집의 다양성을 평가하였으며(Sebiomo et al., 2011; Kim et al., 2012), 토양의 물리·화학적인 지표들과 토양미생물 효소 활성 간의 상관관계를 분석하였다(Adak et al., 2014). 또한 토양 미생물 군집 분석을 이용하여 농업 분야의 미생물의 활용을 검토하였다(Chu et al., 2007; Luo et al., 2015). 그러나, 토양의 건전성 평가 및 이와 관련한 토양미생물에 대한 연구가 주로 농업 분야에 국한되었다(De Gannes et al., 2016). 본 연구에서는 다양한 토양 훼손지의 토양을 채취하여 복원과정의 모니터링을 위한 기본 인자로서 토양의 물리·화학적 특성과 토양 미생물의 생물학적 활성을 분석하여 그 특성을 살펴보았다.
2.1. 훼손토 토양의 시료 채취 장소 및 물리화학적 특성
실험에 사용한 토양은 훼손된 지역의 11개 지점으로, 산불로 피해 입은 토양 3지점, 산사태 발생지 2지점, 개발지 6지점이다. 산불로 훼손된 3지점은 시흥에 위치한 산지로 2018년 4월, 2016년 4월, 그리고 2019년 2월에 각각 1 ha, 0.1 ha과 0.06 ha의 피해를 받았다. 각 지점은 산불 발행 후 각각 5년, 7년, 4년의 시간이 경과하여 그 피해지역이 복원된 상태였다. 그 외, 8지점의 토양은 의왕 1지점, 과천 2지점, 시흥 4지점에서 채취하였다. 각 토양을 채취한 위치의 물리·화학적 특성은 Table 1에 나타냈다. 11개 지점의 토양은 토양공정시험법에 따라 대상지역의 중심이 되는 1개 지점과 주변 4 방위의 1개 지점씩 총 5개 지점을 선정하였다. 토양 표면의 잡초나 이물질을 제거한 후, 토양시료채취기를 이용하여 0 - 15 cm 깊이의 표토를 채취하였고, 모든 시료는 4oC에서 보관되었다. 각 시료명은 임의로 지정된 알파벳과 시료 채취 순서대로 숫자를 지정한 후 표기하였다. 토양의 물리·화학적 특성은 AT분석센터(주)에 의뢰하여 얻은 결과를 이용하였다.
2.2. 효소활성검사
다양하게 훼손된 11지점에서 각각 채취된 토양을 이용하여 탈수소효소와 β-글루코시다아제의 활성을 측정하였다. 탈수소효소 활성 측정은 토양 3 g을 시험관에 넣고 그 후 CaCO3 0.03 g, 3% TTC 1 mL와 멸균수 2.5 mL을 첨가하였다. 광분해를 막기 위해 시험관의 표면을 은박지로 덮고, 37oC 항온기에 넣고 24시간 배양하였다. 배양액에 methanol을 첨가하여, 1시간 동안 흔들어 Ø 110 mm filter paper(ADVANREC, Japan)를 이용하여 반응액을 추출하였다. 추출된 TPF의 양은 485 nm에서 spectrometer (Hach, USA)를 사용하여 측정하였다. β-글루코시다아제(β-glucosidase)의 활성은 QuantiChromTM β-glucosidase Assay Kit(BioAssay systems, USA)를 이용하였다. Assay Buffer 200 μL와 β-NPG 기질 8 μL(final 1.0 mM)을 혼합하여 working solution을 제조하였다. 각 11개 지점에서 채취된 표토를 각각 0.5 g 씩 취한 후 시험관에 넣고 50 mmols/L의 인산 완충용액(pH 7.0) 0.5 mL를 넣고 vortex을 한 후 상등액을 취하여 working solution과 적절히 혼합하여 96 well plate에 분취하였다. 반응액을 포함하는 plate를 Multiskan sky(Thermo scientific, USA)를 이용하여 405 nm에서 흡광도를 측정하고, 아래의 식을 이용하여 계산하였다. 모든 시료의 활성 검사는 3반복 씩 진행하였다.

OD0 = OD 405 nm에서 측정한 0분 때의 시료 값
OD20 = OD 405 nm에서 측정한 20분 때의 시료 값
ODcalbrator = OD 405 nm에서 측정한 20분 때의 값
ODH2O = OD 405 nm에서 측정한 D.W 220 uL를 넣은 셀의 20분 때의 값
2.3. 토양미생물의 탄소원 기질 이용성
미생물의 기질 이용성을 알아보기 위해 EcoPlateTM (Biolog Inc., Hayward, CA)를 사용하였다. Ecoplate는 96 well로 구성된 마이크로 플레이트(microplate) 내에 31종의 다양한 각기 다른 탄소화합물의 기질이 포함된 실험군과 기질이 포함되지 않은 대조군이 3반복으로 되어있으며, 실험군 well에는 다양한 탄소화합물이 얇게 코팅된 상태로 건조되어있다. 31개의 기질은 탄수화물(carbohy- drates, n = 10)계열, 아미노산(amino acids, n = 6)계열, 카르복실산(carboxylic acids, n = 7)계열, 고분자(polymers, n = 4)계열, 페놀화합물(phenolic compounds, n = 2)계열과 아민(amines, n = 2)계열인 6개 그룹으로 되어있다(Sala et al., 2010).
이들 탄소원에 환경 시료 현탁액을 접종하고 배양하면서 시료 내의 미생물군집이 기질을 유일 탄소원으로 이용하여 세균성 호흡이 일어나면 well에 함유되어 있던 tetrazolium dye가 보라색으로 발색되도록 고안되어있다.
시료는 11 지점에서 채취한 표토(0 - 30 cm)를 이용하였다. 토양 시료와 멸균수를 1:10(w/w) 비율로 희석한 후 200 rpm으로 30분간 교반한 후 정치시켜 0.45 um syringe filter로 거른 상등액을 Ecoplate(Biolog, USA)의 각각의 well에 100 μL씩 접종하였다. 이후, 접종된 Ecoplate를 상온에서 120시간 동안 배양하면서 24시간마다 각 well에 배양된 시료의 색을 측정하였다. 배양액의 색 변화는 Mul- tiskan sky(thermo scientific, USA)을 이용하여 595 nm 파장에서 측정하였고 측정된 값을 이용하여 average well color development(AWCD)를 계산하였다. 계산식은 다음과 같다(Garland and Mills, 1991). AWCD는 미생물 군집의 일반적인 대사활성도의 잠재성을 알려주며, EcoPlateTM의 총 생물활성도를 의미하게 된다(Kenarova et al., 2014).

의미 생태학에서 종다양성을 나타내는 지수인 Shanon index 값은 다음과 같은 식으로 계산하였다.

2.4.통계분석
다양하게 훼손된 11개 지점에 대해서 탈수소효소, 베타글루코시다아제, AWCD, Shannon index, 모래(s1), 미사(s2), 점토(s3), 유기물함량(ORGA) 등, 총 8개 변수(차원)에 대해 PCA(Principal Components Analysis) 분석을 하였다(R version 4.0.2). 훼손지 유형을 산불 훼손지를 기준으로 산사태지역과 개발 훼손지를 두 개의 집단으로 각 생물학적 활성의 차이를 알아보기 위하여 유의성 분석을 수행하였다. 유의 확률의 분석 결과 *p<0.05, **p<0.01, ***p<0.005으로 나타내어 집단 간의 차이의 유의수준을 해석하였다.
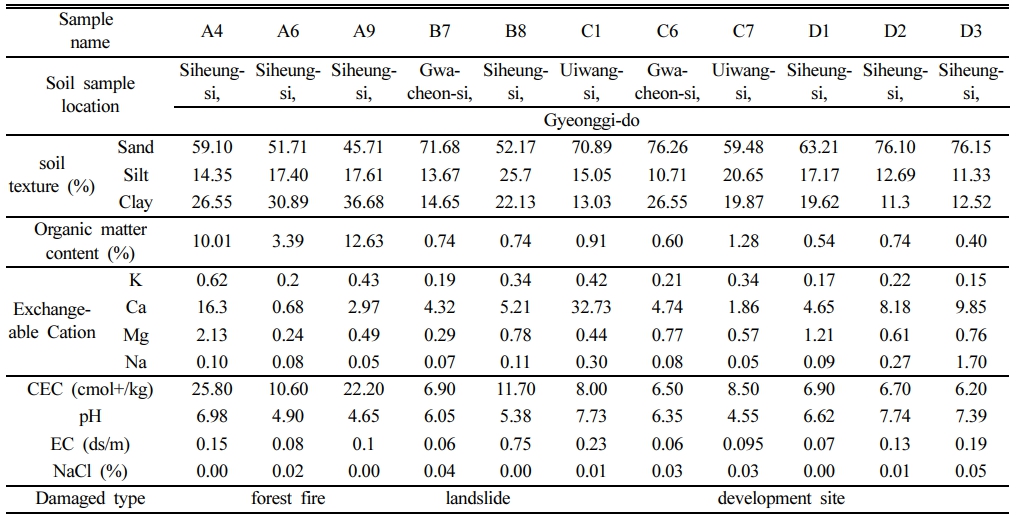
3.1. 훼손토의 효소 활성 비교
다양하게 훼손된 11곳의 지점은 경기도에 위치한 시흥, 과천, 의왕 등지에서 채취하였으며, 토성 중에는 모래의 함량이 많았다. 토양 미생물의 효소 활성은 탈수소효소와 β-글루코시다아제를 측정하였는데, 이러한 토양 효소의 활성은 pH, 토양 유기물 함량, 토양 수분, 습도, 온도, 토양 깊이, 근계(root system), 근권 환경(rhizosphere environ- ment) 및 C-cycle에 영향을 미치는 것으로 알려져 있다(Adetunji et al., 2017; Bergstrom et al., 1998; De Almeida et al., 2015). 탈수소효소는 유기화합물의 호흡에 필요한 산화 환원반응을 촉매하는 세포 내 효소이다. 11개 지점의 토양 시료에 대한 탈수소효소 활성을 측정한 결과를 Fig. 1에 나타내었다. A 지점은 산불에 의해 손상된 산림이고, B 지점은 산사태 발생지역, C와 D 지점은 개발지역으로 구분하였다. A4 지점은 18년도 4월에 1ha가 훼손되었고, A6 지점은 16년도 4월에 0.1ha가 쓰레기소각으로 인해 훼손되었으며, A9 지점은 19년도 2월에 0.06 ha 가량이 농산폐기물소각으로 인한 산불이 발생된 지점이었다.
화재가 발생되었던 A4, A6, 그리고 A9 지점의 탈수소효소 활성은 382.5–937.9μg TPF g-1 24h-1으로 다른 지역에 비해 매우 높은 값을 보여주었다. 산에 화재가 발생하면 표토층의 생태계가 파괴되며, 토양의 물리, 화학, 생물학적 특성이 달라진다. 산불로 훼손된 표토층의 효소 활성에 대한 연구 결과는 연구자들에 따라 화재 기간, 산불 진화 시간, 면적, 회복 기간에 따라 상반된 결과가 보고되었다. Jung et al.(2005)의 연구에서는 산불에 영향을 받지 않은 표토와 산불로 피해를 받고 자연적으로 복원된 표토 간의 탈수소효소 결과, 정상 표토의 탈수소효소 값이 높았다. 유사한 결과를 보이는 또 다른 연구에서도 산불이 발생하기 1년 전의 표토층과 산불이 발생한 시점에서 1년 후에 표토층의 탈수소효소를 비교한 결과, 산불이 발생하기 전의 표토의 탈수소효소 값이 높았다(Memoli et al, 2020). 반면에, 두 산림을 대상으로, 산불로 피해를 입은 토양과 그렇지 않은 토양의 탈수소효소를 비교했을 때, 산불로 피해를 입은 토양의 탈수소효소가 높은 결과를 보이는 연구도 있었다(Singh et al., 2021). 그들의 이전 연구에서도 두 산림을 대상으로, 산불이 발생한 시점으로부터 1, 3, 8, 그리고 13개월 마다 표토층을 대상으로 산불로 피해를 입은 토양과 그렇지 않은 토양의 탈수소효소를 비교한 결과, 산불로 화재를 입은 토양의 탈수소효소가 높았다(Singh et al., 2017). Table 1의 훼손 유형별 토양 특성을 살펴보면, 산불 훼손지는 유기물 함량이 기타 훼손지에 비하여 10배에서 17배 가량 높게 측정되었다.
인위적인 토양 훼손의 유형 중 하나인 산사태 발생지인 B7과 B8 지점에서는 12.61–22.54μg TPF g-1 24h-1 값을 나타내었다. 또한, 개발지역인 C와 D 지점에서는 7.47–51.61μg TPF g-1 24h-1의 다양한 농도 값을 보여주는데, D2 지점은 51.61μg TPF g-1 24h-1로 높은 탈수소효소 활성 값을 보여준다. 이는 D2가 과거에 고물상이 있던 지점으로, 다른 개발지역인 C와 D 지역에 비해 탈수소효소의 농도가 높은 것은 다양한 오염원이 잔류하고 있는 것으로 해석할 수 있다. 일반적으로 탈수소효소 활성은 탄화수소 오염부지, 팜유 공장 폐수부지나 가정폐기물 오염부지와 같이 탄소가 많이 유입되는 부지에서 높은 값이 측정되는 것으로 알려져 있다. 탄화수소, 팜유 잔사물, 생활폐기물 오염 부지들의 표토는 오염되지 않은 표토에 비해서 탈수소효소가 3-7 배 높았음을 보고하는 연구결과가 있었다(Ezirim et al., 2017). Achuba and Pere- tiemo-Clarke(2008)의 연구에서는 폐오일을 농도별로 토양에 접종하여 탈수소효소를 측정한 결과, 대조군에 비하여 오염 토양의 탈수소효소 농도가 최대 2배 높았다. Chinyere et al.(2013)의 연구에서도 pH 변화에 따른 팜유 잔사의 유출수 배출 기점에서 대조군과 비슷하거나 1.3배 정도 높게 나타났다. Hawrot et al.(2005)의 연구에서는 5% 경유로 오염된 사토에서 30일 경과 후, 탈수소효소 값이 대조군에 비해 485% 증가하였다. 산불이 발생되었던 지역도 임업 부산물인 유기물이 연소되는 과정에서 분해됨으로서 유기물 유입이 있는 것과 같은 효과가 있어 효소 활성이 높아진 것으로 볼 수 있다. 산불 훼손지를 기준으로 기타 훼손지의 탈수소효소 활성의 유의 확률(p)을 비교한 결과 훼손 유형별 차이가 있었다(*p<0.05).
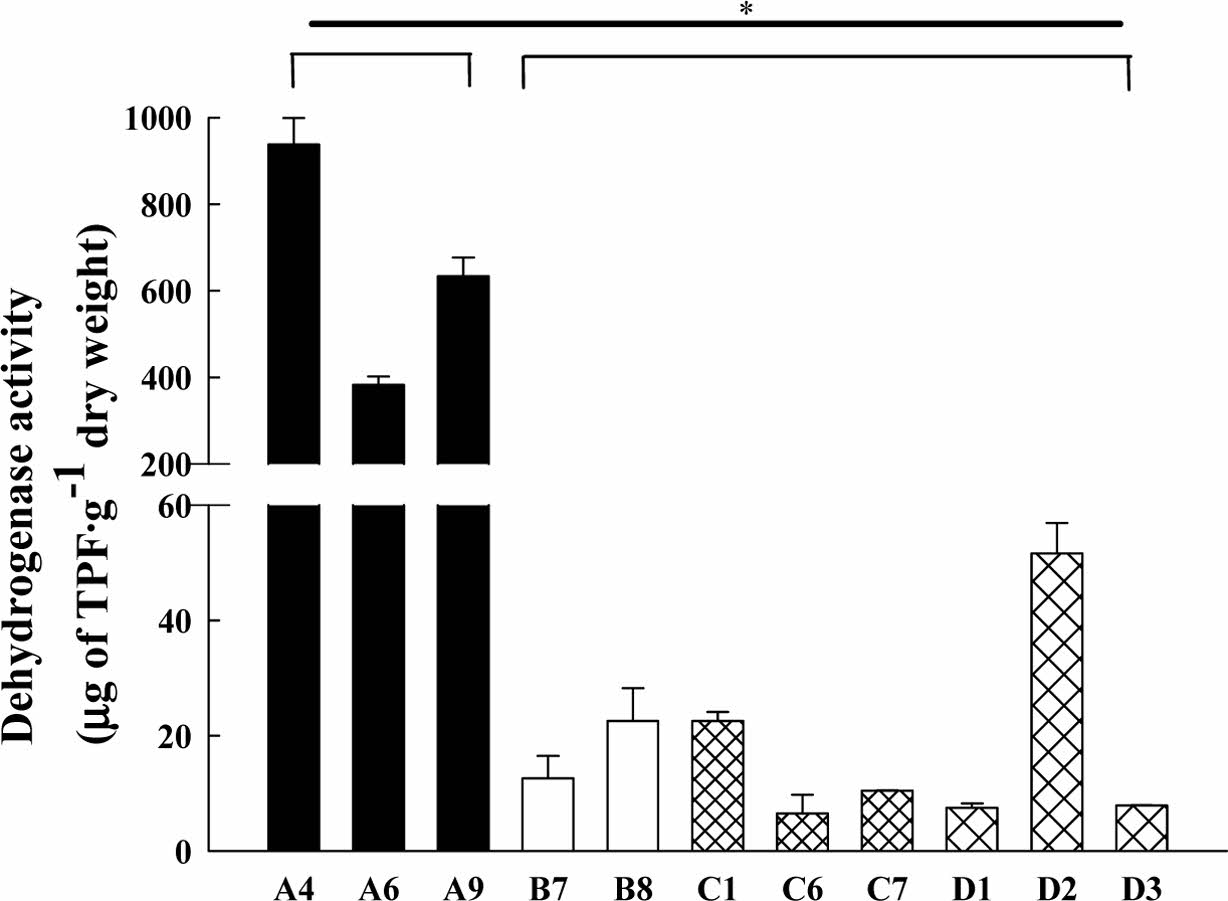
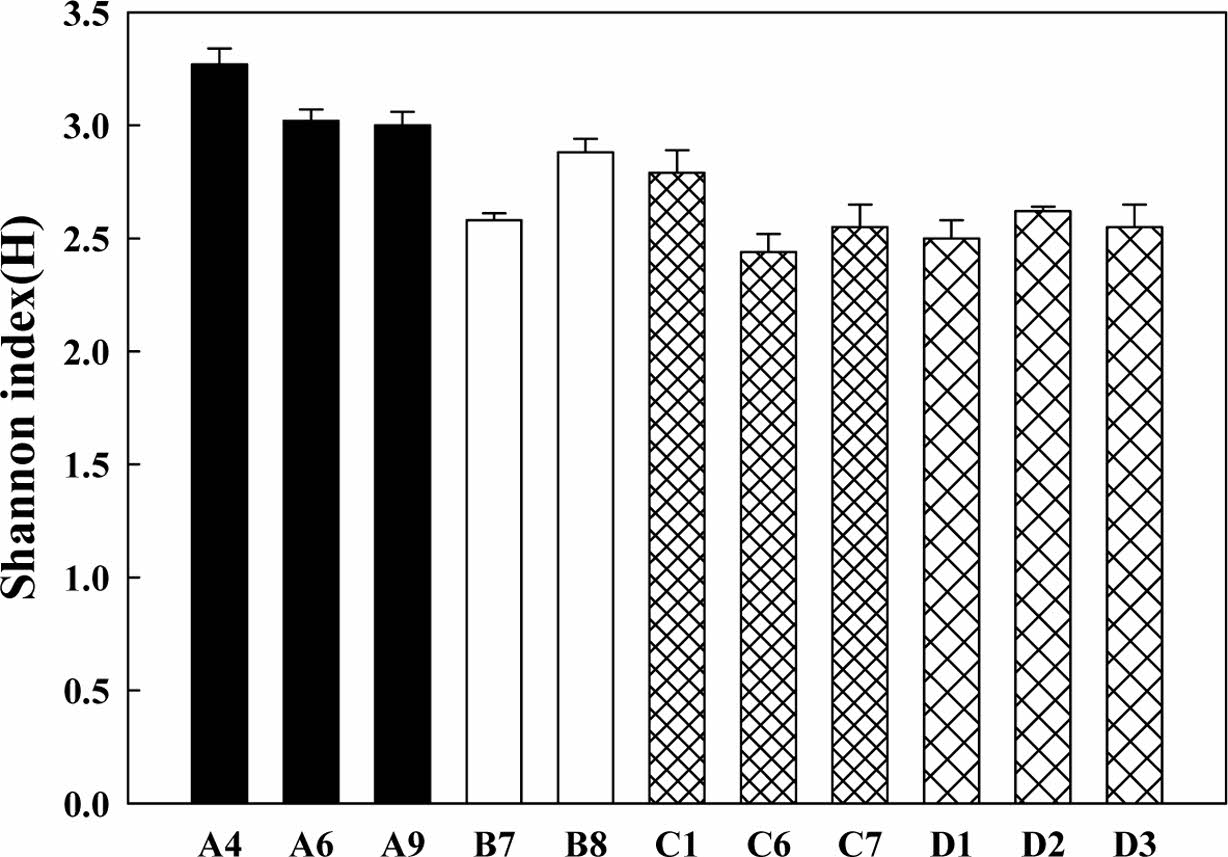
11개 지점에서 채취한 토양미생물의 β-글루코시다아제 활성을 측정한 결과를 Fig. 2에 나타내었다. 산불로 훼손되었던 A 지점이 다른 지점에 비해 2.9–14.5배 높게 나타났다. 섬유소 분해효소 중의 하나로서 저분자의 수용성 섬유소인 셀로비오스, 셀로 올리고당 등을 분해하여 포도당을 생성하는 효소인 β-글루코시다아제는 토양 시료 채취 시기가 산불이 발생한 후 4–7년이 경과된 시점에, 다른 지점보다 아주 높게 나타났다. 즉, 적은 면적에서 화재가 발생되었으나 섬유소로 이루어진 낙엽류가 연소되는 과정에서 수용성 섬유소 성분이 많아지고, 그 지역에 섬유소 분해 효소 활성이 높은 미생물 군집이 우점종을 이룬 것으로 보여진다. 토양미생물의 활성이 높아지면서, 식물의 생장에 유리한 환경이 조성되어 산불로 인한 훼손 후 생태계가 회복되는 과정이라고 해석된다. Singh et al. (2017)의 연구에서는 두 산림을 대상으로, 산불이 발생한 시점으로부터 1, 3, 8, 그리고 13개월 마다 표토층을 대상으로 산불로 피해를 입은 토양과 그렇지 않은 토양의 β-글루코시다아제를 비교하였을 때, 산불로 화재를 입은 토양의 β-글루코시다아제 값이 더 높음을 보여주었다. Oh et al.(2008)의 연구에서는 산불이 발생한 시점으로부터 1, 3, 5, 8, 그리고 10개월마다 표토층을 대상으로 산불로 피해가 있는 토양과 그렇지 않은 토양의 β-글루코시다아제의 활성을 비교한 결과, 산불로 피해가 있는 토양의 -글루코시다아제 활성이 대조토양보다 높지만, 화재 발생 1개월 때의 값이 가장 차이가 많이 나타났고, 점차 시간이 흐를수록 대조토양과 산불로 피해가 있는 토양의 β-글루코시다아제 농도 값이 비슷해지는 것으로 보인다. 일반적으로 산불 발생에 의해 변형된 이화학적 특성은 2-3년 이내에 산불 이전의 상태로 회복된다고 보고되었다(Oh et al., 2008). 이처럼 A지역은 산불이 발생한지, 4-7년 정도 지난 시점이라 산불 발생 이전의 상태로 회복되었다고 해석할 수 있다. B, C, D 지점 중, D 지점들의 효소 활성은 7.91-12.69μmole PNP/g dry weight로 다른 지역보다 높게 나타났다. D1 지점은 야산에 위치하였고, D2는 과거에 고물상이 있던 부지였고 현재 나대지로 남아있으며, D3도 나대지이다. D 지역들은 개발 행위가 개시되기 전 방치되면서 다양한 식생이 자라고 있어 섬유소가 풍부한 환경으로 유지되어 오면서 토양 내 미생물의 섬유소분해활성이 높아진 것으로 보여진다. 산불 훼손지를 기준으로 기타 훼손지의 β-글루코시다아제 활성의 유의 확률(p)을 비교한 결과 훼손 유형별 차이가 있었다(*p<0.05).
토양의 훼손 유형별 토양미생물의 탈수소효소 및 β-글루코시다아제 활성을 비교한 결과, 산불 훼손지의 효소 활성이 기타 훼손지에 비하여 높게 나타났다(Fig. 1). 산불은 섬유소를 분해시켜 토양미생물에 탄소원과 에너지원을 제공함으로서 효소 활성이 매우 높아지는 효과를 보여주며, 이는 훼손토양의 생물학적 활성을 높여주고 복원을 촉진시켜 줄 것으로 사료된다.
3.2. 훼손토 토양미생물의 탄소원 기질 이용성 비교
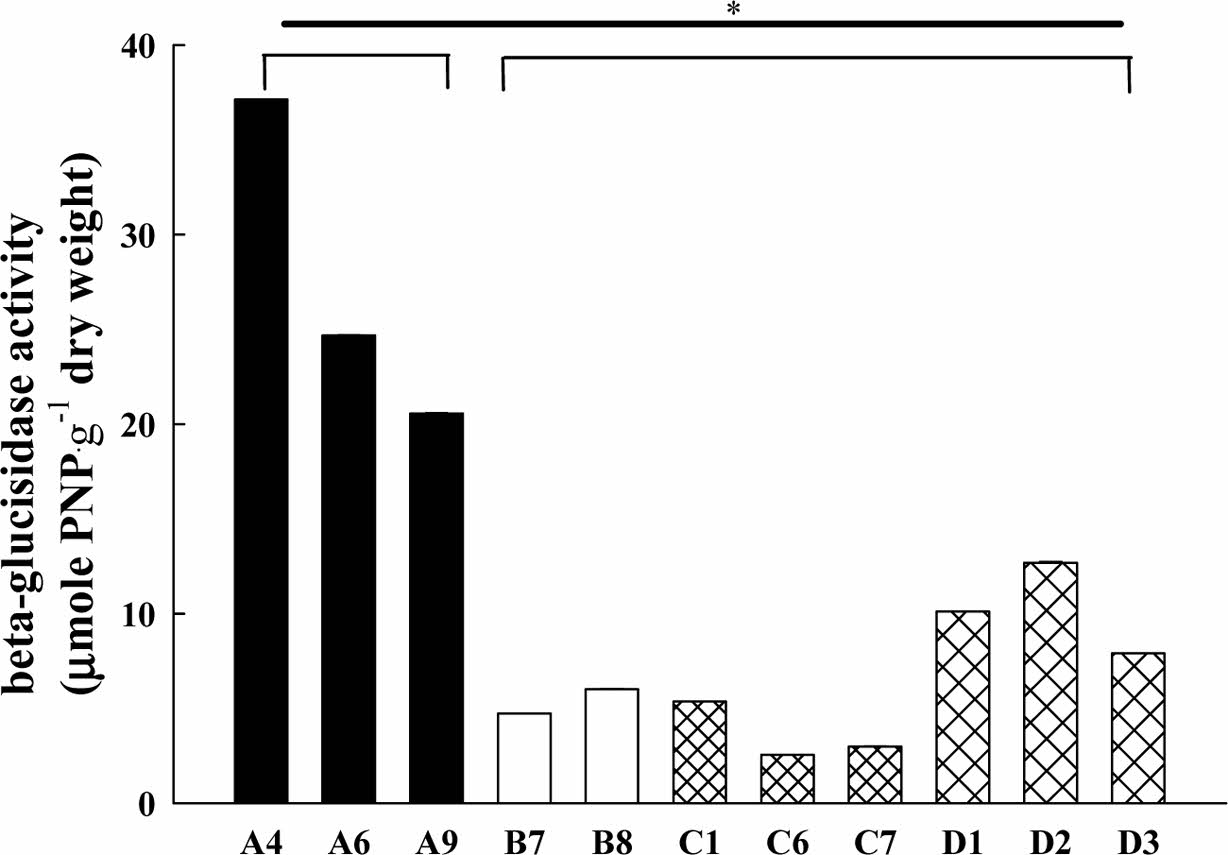
산불, 산사태 및 도시 개발 등으로 훼손된 토양의 생물학적 특성을 알아보기 위하여 토양미생물군집의 생리학적 프로파일링(CLPP, community-level physiological profiling)을 수행하였다. AWCD 값은 토양환경에서 미생물의 총 대사활성도를 알려주는 지표이며, 토양 미생물군집의 대사 프로파일링을 하게 해준다. 이 과정을 통하여 토양 미생물군집이 오랜 시간 동안 특정 공간에서 어떻게 적응하였는지 파악할 수 있다. 이러한 생리학적 프로파일링 방법 중의 하나가 탄소원의 이용 패턴을 알아보는 것인데, 본 연구에서는 EcoPlate에서 제공하는 31가지 기질에 대한 미생물의 이용도를 알아보았다. 각기 다른 토양 시료의 혼탁액을 EcoPlate에 접종하여 120시간 동안 배양하며 얻은 결과를 평균 구획 발색도(Average Well Color De- veolpment, AWCD)로 계산하였고, Fig. 3에 나타내었다. 분석 결과, 산불 지역이었던 A 지역 미생물 군집이 높은 기질이용도 값을 보였다. 다만, 탈수소효소 및 β-글루코시다아제 효소활성도에서는 A 지역이 월등히 높은 값을 보였던 것과 달리, B 지역과 C 지역 미생물군집의 기질이용도 값은 유사하거나 다소 낮은 결과를 보였다.
Weng et al.(2021)의 연구에서는 국립자연보호구역의 3종류 식생에서 AWCD를 측정한 결과, 1.12–1.43 범위를 나타냈다. An et al.(2014)의 연구에서 옥수수를 재배하는 밭 토양에서 AWCD는 대략 0.2 정도의 수치로 보여주지만, 다양한 퇴비를 첨가하면 0.2–1.6 정도의 값들로 올라갔다. An et al.(2015)의 연구에서의 유기농경지 토양의 AWCD 값은 0.81–0.98 범위를 나타냈다. Hong et al.(2011)의 연구에서는 유류로 오염된 토양의 AWCD 값은 1.7이었고, 다양한 토양미생물 복원제를 첨가하면 1.79–2.35로 증가하는 것으로 나타났다. 본연구의 AWCD 값은 유기물이 고갈된 밭토양에 비하여는 낮지 않은 값이나, 인위적 유기물 투입과 오염이 진행된 토양보다 낮은 범위로 보여진다. 반면, A 지역의 값은 유기농경지 지역과 유사하여 화재로 인한 유기물의 분해가 비료투입과 유사한 효과를 보여주는 것으로 생각된다.
일반적으로 다양한 환경에서의 토양미생물의 기질 이용도 분석은 중금속이나, 화학물질로 오염된 토양이나 염도 및 온도 등이 극단적 환경에서의 미생물 군집에서 많은 연구가 이루어졌다. Yang et al.(2022)의 연구에서도 염도-알칼리도가 높아질수록 대사활성도 및 기능적 다양성을 저하시킨다고 보고하였으며, 특히 아미노산 계열의 이용도가 저하됨을 보고하였다. 이와 관련하여 Gemmatimonas, Sphingomonas, Bradyrhizobium 등의 중요 세균 분류군이 심각하게 감소함을 보고하였다. 납으로 오염된 산림지역의 표토와 심토에서의 미생물 활성도 및 기질이용도를 측정한 Borgulat et al.(2021)의 연구에서도 표토의 납 농도가 미생물 활성을 저해하는 것으로 나타났다(Borgulat et al., 2021). Castro et al.(2010)은 기후변화에 의한 환경변화가 미생물 군집의 조성과 대사활성도에 미치는 영향을 알아보기 위하여 4년간 인공적 가뭄을 조성하며 외래식물과 자생식물이 있는 토양을 비교 실험하였다. 강우 시 온도의 증가는 토양의 수분보유력과 불안정한 탄소를 감소시켜주는 것으로 나타났다. 이는 외래식물이 토양의 미세기후를 변화시키고 쉽게 분해되는 고품질의 낙엽을 제공하는 과정에서 미생물 탄소순환에 영향을 주고, 이는 외래식물이 있는 토양에서 발견되는 미생물 분류군과 탄소 대사 유전자의 결과로 알 수 있었다. 즉, 토양미생물이 가뭄과 외래식물종에 반응하면서 잠재적으로 더 작고 취약한 미생물 바이오매스 탄소 저장소를 생산함으로서 생태계의 탄소저장에 영향을 미친다고 보고하였다. De Gannes et al.(2016)의 연구에서 천연숲과 농장(plantation)으로 전환된 숲의 종다양성 지수의 하나인 종 풍부도(species richness) 또는 균등도(eveness)를 비교한 결과 큰 영향이 없었으나, 다변량분석 결과 미생물군집 구성에는 중요한 영향을 미쳤음을 보고하였다. 단변량분석 결과로는 천연숲과에는 세균 문(phyla)이 농장에는 곰팡이 문(phyla)이 각각 풍부하여 종다양성에는 큰 영향이 없으나 특정 균류의 군집에 영향을 미치는 결과 등을 보고하였다.
본 연구 결과, 미생물 군집의 기질 이용도 역시 산불 훼손지가 기타 훼손지에 비하여 높게 나타났다(Fig. 3). 종 다양성 지수인 Shannon index(H)의 값도 AWCD의 값과 유사한 결과를 보여주었다(Fig. 4). 산불 훼손지를 기준으로 기타 훼손지의 기질 이용도와 종다양성 지수의 유의 확률(p)을 비교한 결과 훼손 유형별 차이가 없었다.
3.3. 통계분석
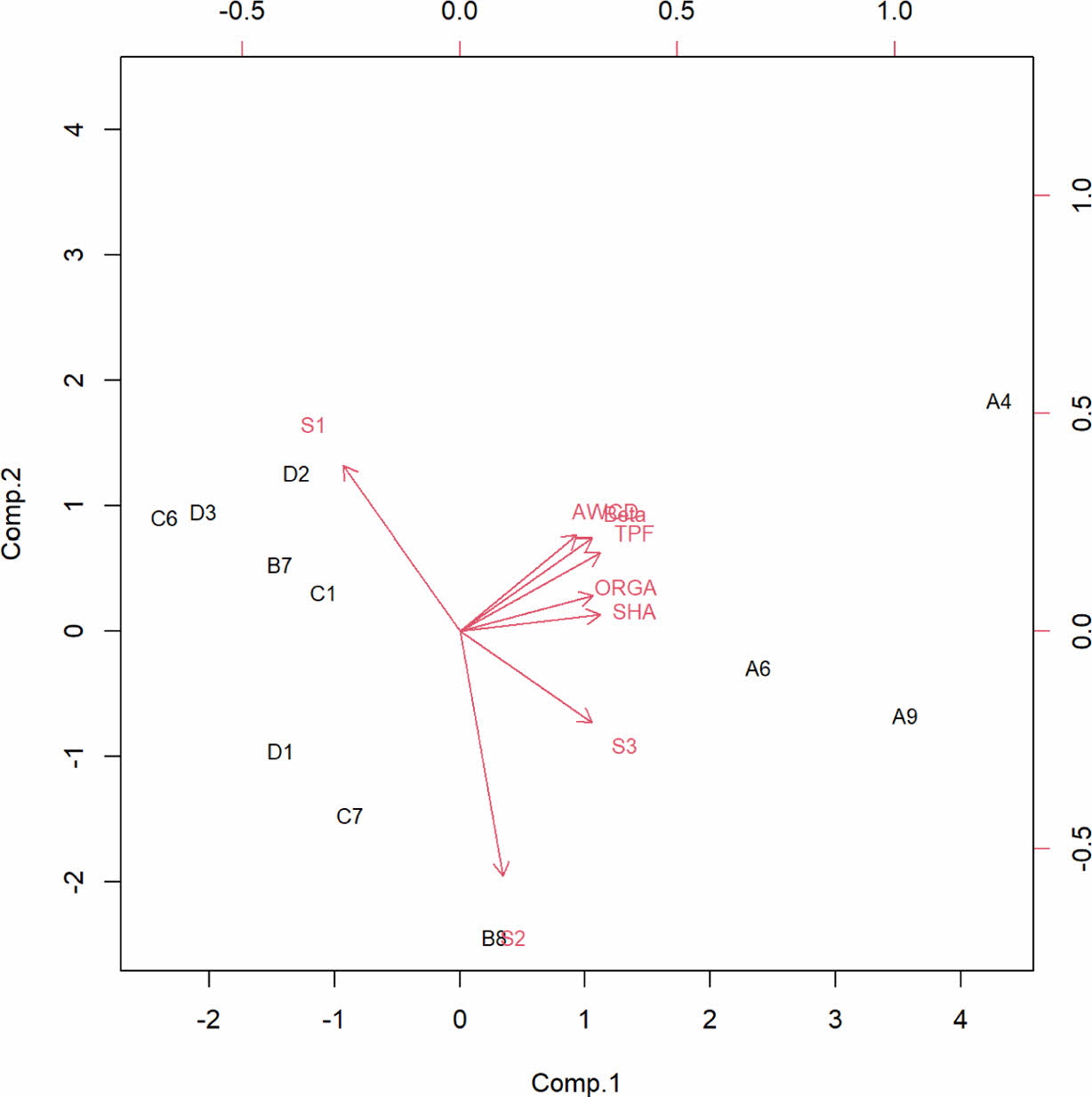
훼손된 각 토양 지점에 대하여 8개 변수를 이용한 주성분 분석 결과, 모래(s1), 미사(s2), 점토(s3) 등의 토성 성분이 각각 독립변수로 작용하였다(Fig. 5). 특히나 모래(s1)와 점토(s3)는 반대적 속성을 나타낸다, 그 외 다른 변수들은 주로 생물학적 활성을 측정한 결과들로 한 그룹을 형성하고 있음을 보여주었다. Fig. 5에서 변수들이 한 방향을 보여주는 그룹은 산불로 훼손되었던 지역들(A4, A6, A9)과 연관이 있는 것으로 나타났으며, 산사태, 개발 및 방치 지역과는 정반대로 분포되는 것을 보여주었다. 즉, 본 연구에서 분석한 훼손지 유형 중 산불 훼손지역의 물리·화학적 변수 중 유기물 함량이 미생물 활성에 깊은 상관성을 보여주고 있다.
Yoon et al.(2022)의 연구에 따르면, 국내 고랭지 밭 토양의 토양 특성 간 상관관계 분석을 통해, pH는 치환성 양이온인 Ca과 고도로 유의한 상관관계를 가진다고 하였다. 이는 석회처리에 따라 Ca 함량이 증가하고 토양 중 pH가 증가하기 때문으로 보았다. 유기물은 pH를 제외한 모든 토양특성과 상관성이 있는 것으로 나타났다. 특히 T-N과 CEC의 경우 상관성이 0.938과 0.920으로 매우 높아 유기물 영향이 큰 것임을 알 수 있었다. 본 연구에서는 토성과 유기물 분석만이 진행되었으나, 유기물과 기타 나머지 화학적 특성의 상관성이 밝혀져 있으므로, 유기물함량이 높은 A지역이 미생물의 활성에 유리한 지역임을 알 수 있다. Lee et al.(2023)의 연구에서도 유기물 함량과 탈수소효소의 고도의 상관관계(p<0.001)를 보였고, 유기물 함량이 높아 유기물에서 수소를 떼어내는 탈수소효소 활성도가 높아진 것으로 언급하였다. 점토광물은 극미립질의 특성과 결정구조적 특징으로 인해 양이온교환능력(CEC, cation exchange capacity)과 비표면적이 높아서 물리화학적으로 점착력 및 화학적 반응성이 매우 높다(Lee et al., 2023). 따라서, 본 연구에서 생물학적 활성도가 모래와 미사보다는 점토질과의 관계에 보다 근접한 것은 위의 화학적 반응성과 상관관계가 높기 때문으로 사료된다. Lee et al.(2023)의 연구에서도 입단 안정성을 판단하는 내수성 입단화율은 유기물, T-N, CEC, 미사와 고도로 유의한 양의 상관관계(p<0.001)를 나타냈고, 용적밀도, 모래와는 고도로 유의한 음의 상관관계(p<0.001)를 보였다. 유기물과 내수성 입단화율간의 상관성이 높은 것은 유기물의 입단형성 효과 때문이며, 이에 따라 유기물과 상관성이 높았던 T-N, CEC 역시 내수성 입단화율과 고도로 유의한 상관관계에 있는 것으로 분석되었다. 그리고 물리적 특성인 용적밀도가 내수성 입단화율과 높은 음의 상관관계에 있는 것은 입단형성에 따라 공극률이 증가하면서 용적밀도가 감소하기 때문이다. 본 연구에서도 이들의 연구처럼 생물학적 활성인자와 모래의 상관관계가 매우 낮은 결과를 보였다.
|
Fig. 1 The dehydrogenase activities of soil microbial communities in various damaged soil (* p < 0.05) |
|
Fig. 2 The β-glucosidase activities of soil microbial communities in various damaged soil (* p < 0.05). |
|
Fig. 3 The Average well color development of soil microbial communities in various damaged soil. |
|
Fig. 4 The Shannon index (H) of soil microbial communities in various damaged soil. |
|
Fig. 5 The Shannon index (H) of soil microbial communities in various damaged soil. |
국내의 다양한 유형으로 훼손된 토양에 대한 물리·화학적 특성과 생물학적 활성을 비교하였다. 산불로 훼손된 지역은 다른 유형과 구별되는 높은 미생물 활성과 높은 탄소원 기질 이용도를 보이는 것으로 나타났다. 이는 산불에 의해 유기물이 분해되어 생물학적 활성이 높아지고, 섬유소 분해를 촉진하여 토양 생태계의 회복을 촉진할 수 있음을 시사한다.
주성분 분석 결과, 토양 성분과 미생물 활성이 서로 다른 그룹을 형성하는 것은 훼손 유형에 따라 달라진다. 산불로 훼손된 지역에서는 유기물 함량이 토양미생물에 의한 생물학적 활성과 관련한 중요한 역할을 하는 것으로 나타났다. 이와 더불어, 모래와 점토 등의 물리·화학적 변수는 미생물 활성보다 다른 화학적 특성과의 상관성이 더 높게 나타났다. 특히, 유기물 함량은 pH를 제외한 대부분의 토양 특성과 상관성을 보이며 높은 유기물 함량은 유기물에서 수소를 떼어내는 탈수소효소 활성을 높이는 것으로 나타났다. 또한, 점토광물은 극미립질의 특성과 결정 구조적 특징으로 물리·화학적 반응성이 높은 것으로 분석된다. 따라서, 유기물 함량이 높아질수록 미생물 활성이 증가하고, 내수성 입단화율과의 관련성이 높아진다.
이러한 결과는 토양 훼손에 따른 생태계 변화와 이에 따른 미생물 활성의 변화를 파악하여 토양 생태계의 회복을 위한 적절한 복원전략을 수립하는데 중요한 정보로 활용될 수 있다.
본 결과물은 환경부의 재원으로 한국환경산업기술원의 표토 보전 관리 기술개발사업의 지원을 받아 연구되었습니다(과제번호: 2020002840001).
- 1. Achuba, F.I. and Peretiemo-Clarke, B.O., 2008, Effect of spent engine oil on soil catalase and dehydrogenase activities, Int. Agrophys., 22(1), 1-4.
- 2. Acosta-Martínez, V., Cruz, L., Sotomayor-Ramírez, D., and Pérez-Alegria, L., 2007, Enzyme activities as affected by soil properties and land use in a tropical watershed, Appl. Soil Ecol., 35(1), 35-45.
-
- 3. Adak, T., Singha, A., Kumar, K., Shukla, S.K., Singh, A., and Singh, V.K., 2014, Soil organic carbon, dehydrogenase activity, nutrient availability and leaf nutrient content as affected by organic and inorganic source of nutrient in mango orchard soil, J. Soil Sci. Plant Nutr., 14(2), 394-406.
-
- 4. Adetunji, A.T., Lewu, F.B., Mulidzi, R., and Ncube, B., 2017, The biological activities of ¥â-glucosidase, phosphatase and urease as soil quality indicators: a review, J. Soil Sci. Plant Nutr., 17(3), 794-807.
-
- 5. Alkortal, I., Aizpurua, A., Riga, P., Albizu, I., Amezaga, I., and Garbisu, C., 2003, Soil enzyme activities as biological indicators of soil health, Rev. Environ. Health, 18(1), 65-73.
-
- 6. An, N.H., Lee, S.M., Cho, J.R., Lee, B.M., Shin, J.H., Ok, J.H., and Kim, S.C, 2014, Effects of long-term fertilization on microbial diversity in upland soils estimated by Biolog Ecoplate and DGGE, Korean J. Soil Sci. Fert., 47(6), 451-456.
-
- 7. An, N.H., Ok, J,H., Cho, J.L., Shin, J.H., Nam, H.S., and Kim, S.C., 2015, Effects of organic matter application on soil microbial community in a newly reclaimed soil, Korean J. Org. Agric., 23(4), 767-779.
-
- 8. Aponte, H., Meli, P., Butler, B., Paolini, J., Matus, F., Merino, C., Cornejo, P., and Kuzyakov, Y., 2020, Meta-analysis of heavy metal effects on soil enzyme activities, Sci. Total Environ., 737, 139744.
-
- 9. Aranda, V., Macci, C., Peruzzi, E., and Masciandaro, G., 2015, Biochemical activity and chemical-structural properties of soil organic matter after 17 years of amendments with olive-mill pomace co-compost, J. Environ. Manage., 147, 278-285.
-
- 10. Bergstrom, D.W., Monreal, C.M., and King, D.J., 1998, Sensitivity of soil enzyme activities to conservation practices. Soil Sci. Soc. Am. J., 62(5), 1286-1295.
-
- 11. Borgulat, J., Kukasik, W., Borgulat, A., Nadgorska-Socha, A., and Kandziora-Ciupa, M., 2021, Influence of lead on the activity of soil microorganisms in two Beskidy landscape parks, Environ. Monit. Assess., 193, 839-852.
-
- 12. Castro, H.F., Classen, A.T., Austin, E.E., Norby, R.J., and Schadt, C.W., 2010, Soil microbial community responses to multiple experimental climate change drivers, Appl. Environ. Microbiol., 76(4), 999-1007.
-
- 13. Chinyere, C.G., Iroha, A.E., and Amadike, U.E., 2013, Effect of altering palm oil and cassava mill effluents pH before dumping on dumpsite soils physicochemical parameters and selected enzyme activities, J. Biodivers. Environ. Sci., 3(4), 46-58.
-
- 14. Chu, H., Lin, X., Fujii, T., Morimoto, S., Yagi, K., Hu, J., and Zhang, J., 2007, Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management, Soil. Biol. Biochem., 39(11), 2971-2976.
-
- 15. Chun, H.C., Lee, S., Gong, D.H., Jung, K.Y., Cho, J.Y., Kim, Y.N., and Lee, Y.B., 2021, Difference in biochemical properties of soils with different periods of agricultural practice after conversion from paddies to uplands, Korean J. Soil Sci. Fert., 54(4), 467-477.
-
- 16. De Almeida, R.F., Naves, E.R., and Da Mota, R.P., 2015, Soil quality: Enzymatic activity of soil ¥â-glucosidase, Glob. J. Agric. Res. Rev., 3(2), 146-150.
- 17. De Gannes, V., Bejeke, I., Dipchansingh, D., Wuddivira, M.N., De Cairies, S., Boman, M., and Hickey, W.J., 2016, Microbial community structure and function of soil following ecosystem conversion from native forests to teak plantation forests, Front. Microbiol., 7, 1976.
-
- 18. Ezirim, C.Y., Chikezie, P.C., Iheanacho, K.M., and Nwachukwu, N.R., 2017, Comparative activities of soil enzymes from polluted sites in Egbema, Imo State, Nigeria, J. Pollut. Eff. Cont., 5(1), 185-192.
-
- 19. Garlamd, J.L. and Mills, A.L., 1991, Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level Sole-Carbon-Source utilization, Appl. Environ. Microbiol., 57(8), 2351-2359.
-
- 20. Hawrot, M., Nowak, A., and Klodka D., 2005, Changes of dehydrogenases activity in soils polluted with diesel fuel, Pol. J. Microbiol., 54(1), 49-53.
- 21. Hong, S.H., Lee, S.M., and Lee, E.Y., 2011, Bioremediation efficiency of oil-contaminated soil using microbial agents, Microbiol. Biotechnol. Lett., 39(3), 301-307.
- 22. Hoorman, J.J., Sa, J.C.M., and Reeder, R.C., 2011, The biology of soil compaction (revised & updated), J. of No-till Agric., 9(2), 583-587.
- 23. Joa, J.H., Moon, D.G., Chun, S.J., Kim, C.H., Choi, K.S., Hyun, H.N., and Kang, U.G., 2009, Effect of temperature on soil microbial biomass, enzyme activities, and PLFA content during incubation period of soil treated with organic materials, Korean. J. Soil. Sci. Fert., 42(6), 500-512.
- 24. Jung, Y.R., Song, I.G., and Kim, Y.J., 2005, Basic study on the biological and physicochemical properties of burnt forest soil for the ecological restoration by organic waste, J. KORRA, 13(1), 79-89.
- 25. Kenarova, A., Radeva, G., Traykov, I.. and Boteva, S., 2014, Community level physiological profiles of bacterial communities inhabiting uranium mining impacted sites, Ecotoxocol. Environ. Saf., 100, 226-232.
-
- 26. Kim, M.I. and Choo, C.O., 2023, Analysis of the effect of forest fires on the mineralogical characteristics of soil, J. Eng. Geol., 33(1), 69-83.
- 27. Kim, J.W., Hong, Y.K., Lee, C.R., and Kim, S.C., 2023, Comparison of physicochemical and biological soil properties in organic and conventional upland fields, Korean J. Soil. Sci. Fert., 56(1), 77-89.
-
- 28. Kim, J.S., Kim, J.H., Jeong, H.C., and Lee, E.Y., 2023, A study on the effects of forest fire on the microbial community activity of forest soil according to the type of rock and difference between topsoil and subsoil, J. Eng. Geol., 33(1), 15-25.
- 29. Kim, Y.H., Lim, J.H., An, C.H., Jung, B.K., and Kim, S.D., 2012, Soil microbial community analysis using soil enzyme activities in red pepper field treated microbial agents, J. Appl. Biol. Chem., 55(1), 47-53.
-
- 30. Langer, U. and Günther, T.H., 2001, Effects of alkaline dust deposits from phosphate fertilizer production on microbial biomass and enzyme activities in grassland soils, Env. Pollut., 112(3), 321-327.
-
- 31. Lee S.J., Choo, C.O, Jeong, G.C., and Kim, J.T., 2023, Analysis and comparison of physicochemical characteristics of first-grade soils from ecological and nature map-focused on igneous rock area-, J. Eng. Geol., 33(1), 61-68.
- 32. Luo, P., Han, X., Wang, Y., Han, M., Shi, H., Liu, N., and Bai, H., 2015, Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China, Ann. Microbiol., 65, 533-542.
-
- 33. Mataix-Solera, J., Cerdà, A., Arcenegui, V., Jordán, A., and Zavala, L.M., 2011, Fire effects on soil aggregation: A review, Earth Sci. Rev., 109(1-2), 44-60.
-
- 34. Memoli, V., Panico, S.C., Santorufo, L., Barile, R., Di Natale, G., Di Nunzio, A., Toscanesi, M., Trifuoggi, M., De Marco, A., and Maisto, G., 2020, Do wildfires cause changes in soil quality in the short term?, Int. J. Environ. Res. Public Health, 17(15), 5343.
-
- 35. Muńiz S., Lacarta J., Pata M.P., Jiménez J.J., and Navarro E., 2014, Analysis of the diversity of substrate utilisation of soil bacteria exposed to Cd and earthworm activity using generalised additive models, PLoS One., 9(1), e85057.
-
- 36. Oh, J.H., Lee, S.B., Park, S.E., Lee, Y,B., and Kim, P.J., 2008, Effect of fire on microbial community structure and enzyme activities in forest soil, Korean J. of Environ. Agric., 27(2), 133-138.
-
- 37. Puissant, J., Jones, B., Goodall, T., Mang, D., Blaud, A., Gweon, H.S., Malik, A., Jones, D.L., Clark, I.M., and Hirsch, P.R., and Griffith, R., 2019, The pH optimum of soil exoenzymes adapt to long term changes in soil pH, Soil. Biol. Biochem., 138(9), 107601.
-
- 38. Sebiomo, A., Ogundero, V.W., and Bankole, S.A., 2011, Effect of four herbicides on microbial population, soil organic matter and dehydrogenase activity, Afr. J. Biotechnol., 10(5), 770-778.
- 39. Singh, A.K., Kushwaha, M., Rai, A., and Singh, N., 2017, Changes in soil microbial response across year following a wildfire in tropical dry forest, For. Ecol. Manage., 391, 458-468.
-
- 40. Singh, D., Sharma, P., Kumar, U., Daverey, A., and Arunachalam, K., 2021, Effect of forest fire on soil microbial biomass and enzymatic activity in oak and pine forests of Uttarakhand Himalaya, India, Ecol. Process., 10(29).
-
- 41. Wade, J., Li, C., Vollbracht, K., Hooper, D.G., Wills, S.A., and Margenot, A.J., 2021, Prescribed pH for soil b-glucosidase and phosphomonoesterase do not reflect pH optima, Geoderma, 401(8), 115161.
-
- 42. Weng, X., Li, J., Sui, X., Li, M., Yin, W., Ma, W., Yang, L., and Mu, L., 2021, Soil microbial functional diversity responses to different revegetation types in heilongjiang zhongyangzhan black-billed capercaillie nature reserve, Ann. Microbiol., 71, 26.
-
- 43. Yang, D., Tang, L., Cui, Y., Chen, J., Liu, L., and Guo, C., 2022, Saline-alkali stress reduces soil bacterial community diversity and soil enzyme activities, Ecotoxicology, 31(9), 1356-1368.
-
- 44. Yoon, J.H., Kim, K.H., and Yang, J.E., 2022, Difference in soil biogeochemical properties of agricultural highland by topographical characteristic and soil management, Korean J. Soil Sci. Fert., 55(1), 1-12.
-
- 45. Zhang, Y.S., Kim, D.J., Cho, H.R., Seo, Y.H., Lee, H.S. and Kim, S.S., 2020, Effects of forest fire on the physicochemical properties of top soils of adjacent agricultural land, Korean J. Soil Sci. Fert., 53(2), 200-208.
-
 This Article
This Article
-
2023; 28(5): 68-77
Published on Oct 31, 2023
- 10.7857/JSGE.2023.28.5.068
- Received on Oct 11, 2023
- Revised on Oct 17, 2023
- Accepted on Oct 24, 2023
Services
Shared
Correspondence to
- Eun Young Lee
-
4The University of Suwon, Department of Environmental and Energy Engineering, Professor, Korea
- E-mail: ley@suwon.ac.kr